When plants get stressed it tends to slow them down. Drought stress – one of the most common causes of losses in crop productivity – can result in wilting, reduce growth, and ultimately, death. And the Climate Crisis means that drought risk is already increasing.
A study by Min May Wong and colleagues, published last year, reveals new key elements that help plants respond to drought, which may have implications for the productivity of plants, in both the good times and the bad.
“Research is definitely an uncertain process and very challenging as the word indicates for, re-and-search. As a PhD student, usually we have to do trial and error many times to get publishable results that could be reproducible. Throughout the process, I was lucky to have very supportive lab-mates and seniors to guide me during the re-search process. Active discussion with supervisor and lab-mates made this study possible.”
Min May Wong, first author of the study
When faced with drought, plants can make certain short-term changes to save water, such as closing the tiny air-holes (stomata) in their leaves that let gasses in and water out, or rolling up their leaves to minimise the leaf surface area. While these measures are important for the plants’ survival, keeping the stress measures activated permanently would also be bad for the plant. Keeping the stomata closed, for example, would prevent the exchange of carbon dioxide and oxygen, effectively shutting down photosynthesis. When it comes to drought stress, and in fact nearly any stress, the key to success is being responsive.
A plant’s ability to respond to drought lies within the cells – in a cascade of molecular reactions that push and pull on the expression of genes and the function of their products, and is ultimately revealed in large-scale changes in plant behaviour growth and development.
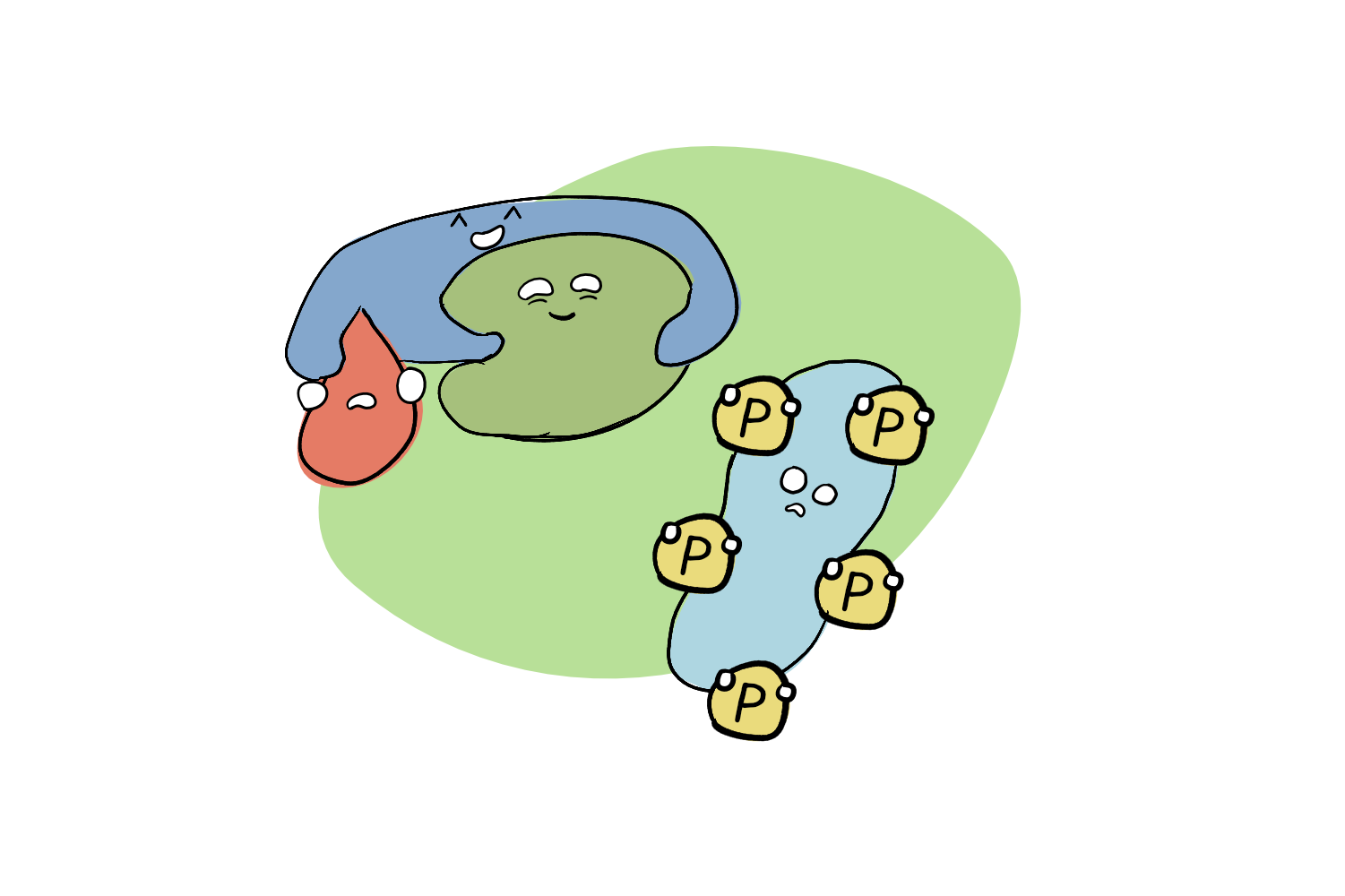
One of the earliest molecular responses to drought involves the accumulation of the hormone ABA (abscisic acid). ABA is a messenger: a molecular alarm that can travel through the plant. Under drought stress, ABA levels increase, which triggers the formation of a complex consisting of the ABA hormone, the receptor protein PYL, and a third protein called PP2C.
PP2C proteins are phosphatases, which means that their day job is to run around the cell removing phosphates from other proteins: a process called dephosphorylation. Removing or adding phosphates to those other proteins is an important way to regulate their activity, effectively turning them ‘on’ and ‘off’ (read more about that here). So when there isn’t a drought, PP2Cs look for certain proteins, and remove their phosphates. But when times get dry, PP2Cs gets ‘hugged’ into ABA-PYL-PP2C complexes, and can no longer do their phosphate-removing job. Suddenly, those other proteins they were targeting get to accumulate a whole lot of phosphates, which changes their own activity!

Although we have a basic understanding of how the process works, we’re still missing a lot of information. For example, we don’t know a lot about which proteins PP2C is regulating. This is a particularly interesting question, because plants don’t make just one type of PP2C, they make a bunch. Which might mean that there are different types of PP2Cs that allow different subtle responses to drought stress.
Min May Wong and colleagues recently investigated a subgroup of PP2Cs known as HAI (Highly ABA Induced), which have been implicated in drought and salt stress, have been shown to affect the plant’s ability to maintain fresh weight under changing osmotic conditions, and may even be involved in bacterial resistance and root development. To understand how HAI proteins might work, Wong and colleagues followed a path of discovery, investigating which proteins are dephosphorylated by HAI, which parts of the protein HAI phosphorylates, and how this leads to the downstream changes that ultimately result in changed plant growth.
First, to discover the targets of HAI (in this case, particularly HAI1), the scientists searched through all the proteins in the cell, looking for those that normally lacked phosphates, but accumulated them when the phosphate-removing HAI1 wasn’t working. For this, they they used a hai1 mutant that couldn’t produce any of the HAI1 protein, but also looked in plants under drought conditions, when HAI1 is known to be locked up in the ABA-PYL-HAI1 complex.

Through this comparison, they were able to find multiple possible targets of HAI1, including several proteins that had already been suggested to be involved in drought stress response.
Unfortunately, correlation does not equal causation: just because some proteins were only de-phosphorylated when HAI1 was active was not proof that HAI1 was doing the dephosphorylation. So, the scientists took it several steps further, focusing specifically on just one of the putative candidate proteins, called AHL. They first showed that AHL and HAI1 could in fact interact within the plant cell, and then undertook an arduous method known as phostag gel analysis, to show that addition of HAI1 to AHL in a test tube could result in removal of the phosphates from AHL.
To verify the phosphoproteomic data and to prove the AHL10 could be directly dephosphorylated by HAI1 [we used the…] wet-lab-based technique phostag gel analysis.
Min May Wong, first author of the study.
This work provides understanding of one important step in the chain of plant growth responses. Drought stress increases ABA, which stimulates the formation of an ABA-PYL-HAI1 complex. This prevents HAI1 from dephosphorylating AHL, which allows AHL to become phosphorylated.
But how does phosphorylated AHL actually make the plant change how it grows under stress? One of the most interesting parts of the study, according to first author Min May Wong, was the finding that phosphorylated AHL10 was able to form small concentrated clusters or foci within the nucleus, while dephosphorylated AHL10 could not. Further research within the study indicated that AHL10 is involved in directly or indirectly changing the expression of growth-related genes, to limit plant growth under drought. Although the identity and composition of the foci remain unknown, the scientists hypothesise that foci formation might required to for AHL10 to do this gene regulation. Wong highlights that future research will involve further dissection of downstream action of AHL10.

Ultimately the authors of the study could suggest multiple potential targets for the phosphatase HAI1, could confirm direct dephosphorylation of AHL10, and could suggest a mechanism by which plants respond to low water by limiting their growth. These findings are extremely important in understanding and securing plant survival in our rapidly warming world.
Interestingly, although the AHL10-controlled growth response is likely an important part of survival under extreme drought, the scientists showed that under moderate drought stress, plants that didn’t have AHL10 activity had a nearly 40% increase in plant mass. This may mean that under certain conditions, the AHL10 activation strategy may be overly conservative, indicating that carefully tuned disruption of AHL10 may even improve plant growth in the right environment.
Reference:
Phosphoproteomics of Arabidopsis Highly ABA-Induced1 identifies AT-Hook–Like10 phosphorylation required for stress growth regulation (2019) PNAS Min May Wong, Govinal Badiger Bhaskara, Tuan-Nan Wen, Wen-Dar Lin, Thao Thi Nguyen, Geeng Loo Chong, Paul E Verslues
We are extremely grateful for Min May Wong’s input on this piece.